

Evoluce kytovců
 |
| Obr. 1 Keporkak - zpívající velryba |
Ačkoliv mohou kytovci připomínat ryby, jsou to ve skutečnosti savci plně přizpůsobení životu ve vodě. Jejich tělo má proudnicový tvar, mají silnou ocasní ploutev, teplotu jim udržuje silná vrstva podkožního tuku, dýchají vzdušný kyslík jako ostatní savci, jejich nosní otvory jsou umístěny nahoře na lebce, jsou schopni echolokace a v neposlední řadě mají sluch vyvinut tak, že jim umožňuje pod vodou vnímat i směr odkud zvuk přichází. Řada těchto charakteristik je činí naprosto unikátními nejen mezi savci, ale vůbec mezi obratlovci. Navíc se jedná o jedny z největších zvířat, která kdy žila na naší planetě. Plejtvák obrovský může dorůst délky až 33 metrů a hmotnosti 180 tun, což ho činní snad největším známým živočichem, který překonává i rozměry největších dinosaurů.
Kytovci (řád Cetacea) se dnes dělí do dvou hlavních skupin - ozubení (podřád Odontoceti) a kosticovci (podřád Mysticeti). Třetí skupinou je vyhynulý podřád Archaeoceti, do kterého patřili společní předci obou dnes žijících skupin. Slovo velryby je vžito jako označení kosticovců, ale používá se i u některých velkých ozubených kytovců. Kytovci jsou blízcí příbuzní sudokopytníků, kam patří také hroch, jehož způsob života je v mnohém stejně unikátní jako kytovců. Podobně jako kytovci tak i hroši rodí pod vodou a stejně tak i pod vodou kojí svá mláďata. Většinu života žijí ve vodě, nicméně jejich adaptace na vodní způsob života nebyla dotažena až do takových detailů jako tomu bylo u kytovců.
Počátek kytovců
 |
 |
| Obr. 2 Andrewsarchus (nahoře) a Indohyus (dole) |
Kytovci vznikli z pozemních savců pravděpodobně z mesonychidů (řád/podřád Mesonychia) nebo sudokopytníků (řád Artiodactyla), což jsou dvě velmi blízce příbuzné skupiny. Meshonychia byla stará skupina prakopytníků, kteří se podobali velkým šelmám i když způsob jejich života byl pravděpodobně poněkud odlišný. Místo drápů měli kopýtka, takže kořist uchvacovali spíše silnými čelistmi, než způsobem jakým to dělají například kočkovité šelmy dnes. Mesonychidi měli výrazně velkou lebku plnou zubů typických pro masožravce. Mezi meshonychidy byli však také býložravci či všežravci. Mezi typické mesonychidy patřil například Mesonyx nebo Andrewsarchus. Obývali Severní Ameriku, Asii a Evropu a žili od spodního paleocénu po svrchní eocén. Mesonichydi Severní Ameriky patřili tehdy k největším dravcům své doby.
Řada vědců se však kloní k názoru, že kytovci vznikli nikoliv z mesonychidů, ale z primitivních sudokopytníků. Předek velryb mohl být také jelenu podobný tvor, který do vody utíkal před predátory. Podobnou strategii dnes zastává kančil, malý praseti podobný tvor, který utíká do vody před orly. Před nedávnem byl v soukromé síbrce geologa Randa Rao objeven primitivní sudokopytník, který je možná nejbližším příbuzným kytovců. Jedná se o rod Indohyus. Indohyus byl asi velikosti lišky, měl silné kosti, což je znak živočichů zdržujících se ve vodě, stavba vnitřního ucha je podoná pritimitním zástupcům z podřádu Archaeocti a v neposlední řadě i jeho zuby vypovídají o tom, že se často pohyboval ve vodě. Indohyus byl nalezen v Kašmíru ve vrstvách starých 55-48 milionů let.
 |
| Obr. 3 Pakicetus |
Jedním z prvních zástupců řádu Cetacea a podřádu Archaeoceti byl Himalayacetus subathuensis. Tento druh byl nalezen ve formaci Subathu v Pakistánu. Bohužel se jedná pouze o nekompletní nález lebky. Nicméně tento druh je důležitý z jiného hlediska. Byl nalezen v mořských usazeninách mezi jinými mořskými živočichy, což vypovídá o tom, že tento živočich žil pravděpodobně v moři nebo moře často navštěvoval. Místo nálezu odpovídá místu, kde se před 50 miliony let rozkládalo mělké teplé moře Tethys. Stáří formace odpovídá asi 53,5 milionu let.
O sto metrů výše ve stejné lokalitě ale v jiné formaci byl nalezen druh Pakicetus inachus. Stáří této části formace Kuldana odpovídá asi 49-50 milionům let. Díky mnohem bohatšímu fosilnímu materiálu víme, že Pakicetus byl primitivním předchůdcem kytovců a přiliš se nelišil od dávných sudokopytníků. Ve skutečnosti spíše připomínal meshonychidy než velryby. Pakicetus společně s dalšími dvěma rody Ichthyolestes a Nalacetus patří do čeledi Pakicetidae. Důkladné studium morfologie těchto prapředků kytovců ukázlo, že již byli částečně adaptováni na vodní prostředíi.
 |
| Obr. 4 Ambulocetus natans |
Ve stejné formaci i když asi o 120 metrů výše byl nalezen další zástupce podřádu Archaeocti. Jedná se o velmi dobře dochovaný exemplář druhu Ambulocetus natans. Ambulocetus byl ještě více přizpůsoben životu ve vodě než tomu bylo u předchozí čeledi Pakicetidae. Ambulocetus je někdy také řazen do samostatné čeledi Ambulocetidae.
Na tuto čeleď navazuje jiná s názvem Remingtonocetidae. Patří sem rody jako Attockicetus, Dalanistes, Remingtonocetus a Andrewsiphius. U všech druhů se jedná o fosilní materiál, který pochází z Indie a Pakistánu. Stáří těchto druhů je řazeno do středního eocénu do období před 40-48 miliony let. Přestože mladší druhy nepatří do vývojové linie vedoucí k recentním druhům, tak se řada vědců domnívá, že předky dnešních kytovců musíme hledat někde uvnitř této čeledi. Remingtonocetidi byli poněkud odlišní od další čeledi Protocetidae. Měli malé oči, dlouhý čenich, ucho velmi podobné dnešních kytovcům a jejich délka nebyla o moc větší než délka dnešních vyder říčních.
Větev s recentním kytovci začíná u čeledi Protocetidae. Jedná se o velmi bohatou čeleď, kterou tvoří tvorové adaptovaní na vodní prostředí avšak stále ještě se všemi plně sformovanými končetinami. Nezapřou však stále své předky, díky nimž mají na všech končetinách kopýtka. Navíc jejich adaptace na vodní prostředí se také projevila na kosti křížové, ktrou tvoří u všech pozemních savců pět spojených obratlů. Zástupci čeledi Protocetidae (podobně jako později kytovci) však tuto kost mají rozdělenu na dvě části. Pánev je však zatím stále pevně připojena k jedné z těchto částí.
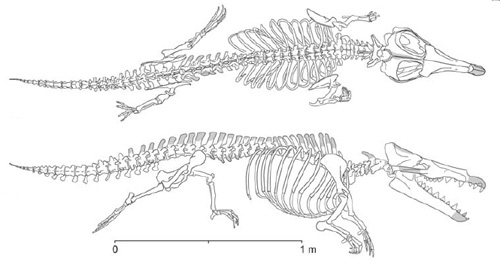 |
| Obr. 5 Maiacetus inuus |
V roce 2009 byl objeven druh Maiacetus inuus. Byly nalezeny dva exempláře z čehož jeden byl pravděpodobně gravidní matkou a měl v těle plod. Poloha plodu v pozici hlavou dolů nasvědčuje tomu, že porod probíhal na souši. Tento druh byl nalezen v horninách starých asi 47,5 milionu let.
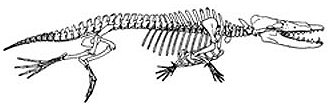 |
 |
| Obr. 6 Rodhocetus kostra a umělecká rekonstrukce |
Jedním z výborně dochovaných exemplářů je Rodhocetus. Byly popsány dva druhy R. kasrani a nedávno objevený druh R. balochistanensis. Rodhocetus byl přizpůsoben vodnímu životu daleko více než Ambulocetus. Jeho vnitřní ucho bylo již velmi podobné dnešním kytovcům a nosní otvor byl posunut od čenichu směrem dozadu. Jeho pohyb pod vodou však byl zcela jistě odlišný od způsobu pohybu dnešních kytovců. Rodhocetus měl silné končetiny a používal je pravděpodobně jako pádla, kdežto dnešní kytovci používají pro pohyb téměř výhradně ocasní ploutev. Tu však Rodhocetus ještě neměl. Jeho pozůstatky pocházejí spíše z hlubokomořských usazenin než z mělkých moří. Byl nalezen ve formaci Domanda v Pakistánu. Této formaci odpovídá stáří asi 46-46,5 milionu let.
Dalším druhem z této bohaté čeledi, který je znám na základě bohatšího fosilního záznamu je Protocetus atavus. Byl objeven ve formaci Mokattam v Egyptě. Stáří lokality náleží do středního eocénu asi 38-48 milionů let. Protocetus byl stejně jako Rodhocetus přizpůsoben životu pod vodou. Jeho střední ucho byla adaptováno pro vodní prostředí, nicméně dle všeho ještě nebyl schopen echolokace. Neidentifikovatelný obratel z Texasu v USA náležící taktéž do středního eocénu byl přisouzen také tomuto rodu. Zařazení je však nejisté a nebylo tomuto exempláři přisouzeno ani druhové jméno.
 |
| Obr. 7 Basilosaurus |
Georgiacetus je opět jedním z čeledi protocetidae, který je znám na základě bohatšího fosilního materiálu. Byl nalezen ve formaci McBean jejíž stáří je odhadováno na 40-41 milionů let. Zástupci této čeledi jsou prvními předchůdci kytovců, kteří se rozšířili po většině mělkých tropických moří světa.
Na tento druh navazuje čeleď basilosaridae, která se dělí na dvě podčeledi - basilosaurinae a dorudontinae. Skupina okolo rodu Basilosaurus z podčeledi basilosaurinae je zvláštní svou specifickou stavbou obratlů. Jejich tvar připomíná spíše obratle plaza. Proto byl první známý zástupce této skupiny pojmenován Basilosaurus, což znamená "královský plaz". Později po prozkoumání celé kostry bylo jasné, že se nejedná o plaza, ale o savce. Jméno však již zůstalo. Basilosaurus se vyznačuje enormně dlouhým tělem s předními končetinami ve tvaru ploutví a zakrnělými zadními končetinami. Právě díky jeho obratlům se paleontologové domnívají, že se jedná o slepou evoluční větev, ze které se žijící kytovci nevyvinuli.
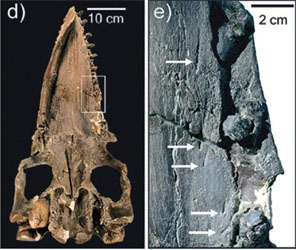
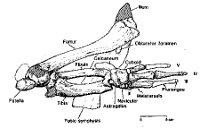 |
| Obr. 8 Zadníkončetina Basilosaura (měřítko v pravo dole znázorňuje 5cm) |
Druhá podčeleď této čeledi má velmi podobné charakteristiky jako basilosauři, nicméně obratle nemají tak protáhlé jako zástupci podčeledi basilosaurinae. Jedním zástupcem této skupiny je Dorudon s výborným fosilním záznamem. Vyznačuje se opět dlouhým protáhlým tělem, rozekladným ocasem, předními končetinami ve tvaru ploutví a malými zakrnělými zadními končetinami. Jejich stavba se podobá té u basilosaura. Tyto malé končetiny už nejsou pevně připojeny k pánvi. Dle celkové stavby kostry lze usuzovat na to, že zástupci této čeledi nebyli tak zdatnými plavci jako jsou dnešní kytovci. Dorudon dosahoval délky asi 5m. Byl nalezen společně s Basilosurem ve formaci Jebel Qatrani oblasti Fayum v Egyptě. Stáří této formace se odhaduje na 33-37 milionů let, což odpovídá svrchnímu eocénu až spodnímu oligocénu.
Mezi množstvím koster dorudona bylo nalezeno i několik mladých jedinců. Někteří z nich měli na lebkách stopy po velkých zubech, které odpovídají zubům basilosaura. Je tedy možné, že dospělí basilosauři lovili mláďata dorudona.
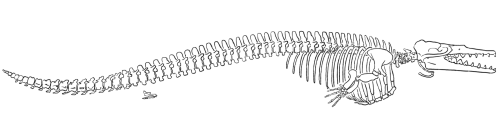 |
| Obr. 9 Dorudon |
Do této čeledi patří další rody jako například Ancalecetus, Pontogeneus, Saghacetus a další. Tyto rody jsou známy z méně kompletního fosilního záznamu a stejně jako druhy popsané výše, byly nalezeny v Egyptě ve vrstvách spadajících do svrchního eocénu až spodního oligocénu.
Ozubení (Odontoceti)
Nyní se dostáváme do spodního oligocénu, kde se cesty kytovců dělí na dvě významné skupiny. Podřád kosticovců a ozubených. Od této chvíle spadají oba podřády do skupiny s názvem Autoceta. Skupina ozubených kytovců je bohatší než skupina kosticovců. Pouze některým zástupcům této skupiny se vžil název velryba. Jako velryby běžně ozančujeme kosatky nebo vorvaně i když s pravými kosticovci nemají nic společného, neboť jsou to právě ozubení kytovci. Dalšími zástupci patřícími do této skupiny jsou známí delfíni, narvali, běluhy, atd.
Podřád ozubených se dále dělí asi do čtyř nadčeledí. Physeteroidea (vorvani a kogie), Hyperoodontoidea (vorvaňovci), Delphinoidea (delfíni, kosatky, sviňuchy, běluhy apod.) a Platanistoidea (delfínovci). Cesty prvních dvou skupin od ostatních se oddělili někdy během spodního oligocénu.
Physeteroidea
 |
| Obr. 10 Vorvaň obrovský |
Skupina se dělí někdy na jednu jindy na dvě čeledi. Jedná se o čeleď Physeteridae (vorvaňovití), která se někdy ještě dělí na čeleď Kogiidae (kogiovití). Vorvaňi jsou typičtí velkou hlavou, která obsahuje tzv. spermacet. Jedná se o voskovitou látku, která nemá nic společného se spermatem. V angličtině se dokonce vžil název "sperm whale". K jakému účelu tento orgán slouží lze pouze spokulovat. Existují tři hypotézy, které se snaží vysvětlit účetl této zvláštní struktury. Údajně se může jednat o orgán, který velrybám umožňuje lépe vyrovnávat tlak ve velkých hloubkách. Nebo se může jednat o "nástroj" pro boj mezi samci. Nebo to může být orgán, který vylepšuje echolokaci.
Nejstarší kytovci z této skupiny pochází až ze středního miocénu. Tedy z období před asi 16 miliony let. O jejich původu tedy mnoho nevíme. Předchůdci vorvaňů měli narozdíl od žijících druhů zuby jak ve spodní tak i v horní čelisti. Dnešní vorvani mají zuby pouze ve spodní čelisti, které zapadají do důlků v čelisti horní. Už v miocénu byli vorvani velkými dravci. Jedním z nejlépe zdokumentovaných předchůdců je Zygophyseter varolai. Jedná se o druh žijící ve svrchním miocénu asi před 7-11,5 miliony let na území dnešní Itálie. Byl nalezen téměř kompletní exemplář, takže máme k dispozici výborný materiál pro studium.
 |
| Obr. 11 Zygophyseter varolai |
 |
| Obr. 12 Kogie |
Dalším vorvaněm ze svrchního miocénu je rod Aulophyseter s třemi druhovými jmény. Tento rod je znám na základě fosilního materiálu ze Severní Ameriky. Dále se skupina dělí na další recentní podčeleď (čeleď). Jedná se o kogiovité (Kogiinae), kam patří kogie. Poměrně neznámé druhy velryb blízce příbuzné vorvaňům. Dnes žijí dva druhy kogie. Jedná se o kogii tuponosou (Kogia breviceps) a kogie Owenovu (Kogia simus). Kogie jsou malí kytovci dosahující délky 2,5-3,5 metru. V pliocénu asi před 2-5 miliony let žil ještě třetí druh K. pusilla. Kromě tohoto rodu známe ještě další vyhynulé druhy patřící do této podčeledi jako Praekogia cedrosensis nebo Scaphokogia cochlearis.
Druhá podčeleď Physeterinae obsahuje řadu miocenních druhů jako byl Placoziphius duboisii, Orycterocetus crocodilinus, Idiophyseter merriami nebo Physeterula dubusii. Z této podčeledi přežil dodnes pouze vorvaň obrovský (Physeter macrocephalus). Ten je také současně největším žijícím zástupcem této nadčeledi.
Hyperoodontoidea
 |
| Obr. 13 Eurhinodelphis |
Ve fosilním záznamu jí předchází čeleď s názvem Rhabdosteidae. Tato čeleď je charakteristická protaženou horní čelistí. Mezi první zástupce této čeledi patří rod Schizodelphis. Objevil se ve spodním miocénu asi před 28 miliony let a přežil až do svrchního miocénu před asi 7,5 miliony let. Byl objeven v západní Evropě a Severní Americe. Později asi před 23 miliony let se objevil další druh této skupiny - Ziphiodelphis abeli. Zhruba ve stejnou dobu se objevuje další zástupce rod Eurhinodelphis. U tohoto rodu bylo popsáno hned několik druhů, kde nejstarší žil v pozdním až středním miocénu a ti nejmladší druhy přežili až na hranici spodního a pozdního pliocénu asi před 2 miliony let.
 |
| Obr. 14 Vorvaňovec Layardův (Mesoplodon layardii) |
Od této skupiny se oddělila právě i nadčeleď vorvaňovců. Jedná se o málo známé tvory, kteří připomínají velké delfíny. Dorůstají úctyhodných délek až kolem deseti metrů. Jsou typičtí dvěma velkými zuby ve spodní čelisti, jejichž tvar a velikost se druh od druhu liší. První zástupci této skupiny se objevili už ve spodním miocénu.
Dnes je popsáno šest rodů a celá řada druhů vorvaňovců. Mezi žijící vorvaňovce patří rody Tasmacetus a Berardius z podčeledi Berardiinae dále pak Indopacetus, Hyperoodon a Mesoplodon z podčeledi Hyperoodontinae a posledním rod je Ziphius z podčeledi Ziphiinae.
Delphinida
 |
| Obr. 15 Delfín |
Do této skupiny patří všichni defínovci (kromě delfínovce ganžského), běluhy, narval, orcela, sviňuchy, kosatky, delfínci, plískavice, kulohlavci, elektry, ferezy a samozřejmě delfíni. Je to opravdu velmi bohatá skupina mořských savců čítající desítky druhů. Jedná se převážně o malé druhy. Výjimku tvoří pouze kostaky, které se vyrovnají velikostí vorvaňům a velrybám. Jednotlivé druhy jsou velmi podobné delfínům tak, jak je známe. Většinou se liší zbarvením a tvarem hlavy. Zatímco delfíni mají malý protáhlý čenich, tak běluhy, kulohlavci, ferezy, elektry, sviňuchy a další mají spíše kulovitý tvar hlavy s menším čenichem. Na druhou stranu delfínovci mají čenich štíhlý a protáhlý.
Jakousi výjimku tvoří narval, který má jeden zub tvarovaný do dlouhého rohu, který ční z horní čelisti. Jedná se většinou o levý řezák. Pravý bývá zakrnělý, ale občas se vyvine i on a narval tak má dva dlouhé kly. Samice tento kel mají také, ale velmi malý ukrytý v dásni. Účel tohoto rohu/klu dosud není přesně znám. Blízkým příbuzným narvalů je také rod Odobenocetops. Tito kytovci měli také kel, jehož růst se vymkl kontrole. Kel však nesměřoval vpřed, ale naopak vzad. Většina jedinců měla kly dva, kde jeden byl větší než ten druhý. U některých druhů se vyvinul pouze jeden kel. Odobenocetops žil ve spodním pliocénu před asi 5,5-3,5 miliony let.
 |
 |
| Obr. 16 Narval a Odobenocetops | |
Platanistoidea
 |
| Obr. 17 Delfínovec ganžský |
Poslední nadčeleď je zastoupena dnes pouze jedním rodem Platanista. Jedná se o delfínovce, který říje v zakalených řekách. Je téměř slepý, a proto pro orientaci využívá echolokaci. Má dlouhý štíhlý čenich, který se směrem ven rozšiřuje a skrývá dlouhé zuby, které jsou patrné i když má delfínovec tlamu zavřenou. Tito říční delfínovci žijí v řekách Indus a Ganga na území Indie, Nepálu, Pákistánu a Bangladéše.
Kosticovci (Mysticeti)
Skupina kosticovců je typická tím, že z jejich horní čelisti místo zubů vyrůstají kostice. Těmi velryby filtrují plankton a malé živočichy, kterými se živí. Kosticovci se dělí asi na čtyři čeledi - Balaenopteridae (plejtvákovití), Balaenidae (velrybovití), Neobalaeonidae (velrybkovití) a Eschrichtiidae (plejtvákovcovití).
 |
| Obr. 18 Janjucetus hunderi |
Z hornin starých asi 23-28 milionů let známe jedny z nejprimitivnějších kosticovců. Jedná se o druh Janjucetus hunderi. Byla nalezena kompletní lebka. Je z ní patrné, že první kosticovci ještě žáné kostice neměli. Junjacetus byl nalezen na území dnešní jižní Austrálie. Na stejném území a ve stejné vrstvě byl nalezen také druh Mammalodon colliveri. Ze západu USA a Kanady známe další pozdně oligocenní rod - Chonecetus. Z Japonska a USA pak známe rod Aetiocetus. Tyto druhy se vyznačují nosním otvorem posazeným hodně dozadu, nicméně ještě ne tak daleko, jako je tomu u dnešních velryb. Nejzajímavější jsou však posledně jmenované dva rody. Jedná se totiž o první ozubené kosticovce, kteří již měli také kostice. Kostice bohužel nefosilizují, ale nechali nám po sobě stopy. U žijících velryb kostice vyrůstají z horní čelisti a jsou zásobeny krevními žilami, po nichž zůstavají v horní čelisti malé kanálky. Právě Aetiocetus měl vedle zubů podobných například těm u druhu Junjacetus, také tyto kanálky v horní čelisti, což svědčí o tom, že měl za života i kostice.
 |
 |
| Obr. 19 Aetiocetus rekonstrukce (vlevo) a lebka s vyznačením kanálků vyživujících kostice (vpravo) | |
Ve středním až svrchním miocénu se od větve směřující k recentním velrybám oddělila čeleď Cetotheriidae. Druhově nerozmanitější rod této čeleď rod Cetotherium žil od spodního miocénu po pliocén (1,8-20,5 milionu let.) na území dnešní USA a Evropy. Nejstarším druhem byl druh C. furlongi a nejmladším pliocenním druhem druh C. crassangulum. Kromě tohoto druhu čeleď obsahovala rody jako například Mixocetus, Metopocetus, Herpetocetus a další. Všechny tyto rody však nepřežili hranici pliocén/pleistocén. Zástupci této čeledi připomínaly malé velryby - délka se pohybovala okolo pěti metrů a váha kolem 1 tuny. Jejich kostice byly krátké narozdíl od dnešních velryb. Fosílie nám také dokládají, že tito malí kytovci byli často loveni velkými předchůdci dnešních žraloků.
 |
| Obr. 20 Cetotherium |
 |
| Obr. 21 Kostice |
Nejstarším fosilním kytvcem, který má již pouze kostice je Eomysticetus. Tento rod byl nalezen v horninách svrchního oligocénu Severní Karolíny v USA. Vrstvy spadající do této formace jsou staré 23-28 milionů let. Dalšími druhy v linii vedoucí k recentním druhů byly např. Diorocetus, Pelocetus, Aglaocetus a další. Jedná se o druhy od spodního miocénu (23 mil. let) až po střední miocén (11,5 mil. let). Tyto druhy již obývaly prakticky celou planetu od USA, přes Evropu, východní Asii až po Jižní Ameriku.
Ve svrchním miocénu se linie štěpí na všechny dodnes žijící čeledi kosticovců. Kosticovci mají místo zubů kostice, kterými filtrují plankton. Jedná se o útvary z keratinu, tedy materiálu, ze kterých jsou nehty, vlasy nebo rohy. Kostice jsou dva paralelní řady dlouhých desek podobných hřebenu. Délka kostic je 0,5-3,5 metru. Nejdelší kostice má velryba gronská (Balaena mysticetus), která také patří mezi největší současné velryby. Rekordmanem mezi velrybami je ovšem plejtvá obrovský (Balaenoptera musculus), který může dosahovat délky až 33 metrů a váhy 180 tun.
 |
 |
| Obr. 22 Plejtvák obrovský a velryba grónská | |
Doklady evoluce
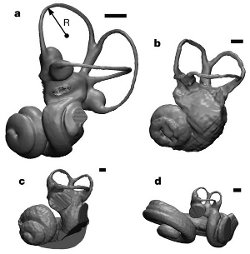 |
| Obr. 23 Redukce labyrintů při vývoji vznitřního ucha. a) Galago moholi - běžný savec b) Ichthyolestes pinfoldi c) Indocetus ramani (částečně rekonstruovaný) d) Tursiops truncatus. Měřítko (černý proužek) je 1mm. |
Původ kytovců je jednou z nejlépe zmapovaných cest evoluce. I přesto, že máme stále řadu mezer, tak náš obraz vývoje kytovců je velmi dobrý a téměř všechny významné fáze máme zmapované díky poměrně dobrému fosilnímu záznamu. Můžeme tak sledovat naprosto unikátní cestu pravěkých kopytníků, kteří se vydali po 300 milionech let zpět do vody.
Nejvýznamnějším dokladem evoluce jsou hlavně unikátní znaky, které kytovci mají. Jedná se mimo jiné o stavbu vnitřního a vnějšícho ucha, která nám ukazuje postupnou změnu od běžného savčího ucha po specializované ucho kytovců. Jedná se hlavně o redukci velikosti labyrintů, která je mezi savci naprosto unikátní. Změny můžeme pozorovat v několika fázích. Od běžného savčího typu, který je reprezntován u čeledi Pakicetidae. Přes jakýsi kompromis, který fungoval dobře jak na souši, tak pod vodou u čeledí Remingtonocetidae a Protocetidae. Až po sluch plně adaptovaný na podvodní život, který již na souši není použitelný. Tuto fázi blízkou stavu, který nacházíme u recentních druhů můžeme vidět u čeledí Basilosauridae a Dorudontidae.
Dalším unikátním znakem je nosní otvor posazený u dnešních kytovců vysoko na hlavě. I tuto změnu nám krásně demonstruje fosilní záznam. Můžeme vidět u pakicetidů normální polohu nosních otvorů vpředu na čenichu. U protocetidů se otvor posouvá dále od špičky čenichu. U basilosauridů a dorudontidů je někde na půli cesty. A jeho přesun dále dozadu můžeme sledovat i u obou hlavních skupin - kosticovců a ozubených.
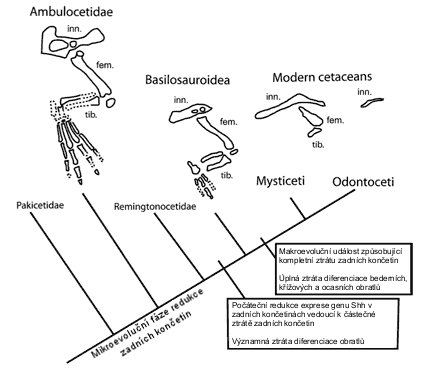 |
| Obr. 24 Postupná ztráta zadních končetin |
 |
|
Obr. 25 Delfín chycený u |
Také ztráta končetin je typická pro kytovce. Bohužel u ztráty končetin nám chybí fosilní záznam, který by ukazovat změnu od dlouhých zadních končetin protocetidů po malé zakrnělé končetiny dorudontidů a basilosauridů. Nicméně dle genetických studií je ztráta končetin, které již nejsou potřeba, poměrně malou a rychlou změnou. Ve skutečnosti delfíni mají plně funkční všechny potřebné geny, které formují zadní končetinu. Změnou prošly pouze regulační geny, které je ovládají. Jedná se hlavně o zkrácenou dobu exprese hegu Shh (Sonic hedgehod). Naštěstí nám malinké zadní končetiny basilosauridů a dorudontidů, které již nejsou vůbec použitelné na souši, ukazují pravý původ těchto pravěkých kytovců.
Původ kytovců z pozemních čtyřnohých tvorů dokládají i dnešní zbytky končetin a pánve u recentních druhů. Kosticovci i ozubení mají v sobě stále ještě zbytky pánve (u kosticovců ještě zbytky velkých kostí zadních končetin - kost stehenní a kost holenní), která již není pevně přichycena k obratlům a slouží pouze jako opora pro pohlavní orgány stejně jako je tomu mimo jiné i u terestriálních savců. Ostatní funkce jako u pozemních savců však pánev již neplní. Navíc občasné atavismy čtyřnohý původ nezapřou. V roce 2006 byl chycen u Japonska dokonce delfín s vyvinutými zadními končetinami.
O čtyřnohém původu nám také vypovídá embryologie, kdy se v počátečních fázích ontogeneze vytváří i pupeny pro zadní končetiny, které jsou pak v pozdějších stádiích vstřebány a dále se nevyvíjí. Stejně jako zuby u kosticovců, které se také objevují během otogeneze, ale během dalšího vývoje mizí.
Závěr
I přesto, že máme v poznání evoluce kytovců bílá místa a ve fosilním záznamu řadu mezer, tak to co máme k dispozici, nám již dává jakýsi ucelený obraz o tom, jak, kde a kdy evoluce kytovců probíhala. Nejvíce jsme se dozvěděli o evoluci velryb během posledních asi 20ti let, kdy byla nalezena řada přechodných forem ve fosilním záznamu. Jestli dosavadní tempo výzkumu evoluce kytovců bude pokračovat, tak se máme v blízké budoucnosti na co těšit.